INTRODUCTIONVibrio cholerae is a microorganism that causes acute diarrheal diseases and cholera, one of the leading causes of global morbidity and mortality, especially in children under five years old. It is present in many regions and has been isolated from diverse sources such as water, soil and food. Surveillance of this microorganism in Cuba from 1985 through June 1997 showed circulation of non-epidemic non-O1/non-O139 serogroups, but surveillance continued to identify distribution of V. cholerae serotypes and serogroups in the different geographic regions of the country during the following years, due to the risk of introducing cholera-causing serogroups that provoked cholera epidemics in other countries of the region.
OBJECTIVES Describe the temporal‒spatial distribution of serogroups and serotypes of V. cholerae in Cuba.
METHODS A cross-sectional study was conducted that included isolates from passive surveillance of V. cholerae in 16 hygiene and epidemiology centers throughout Cuba from July 1997 through December 2019, submitted to the National Reference Laboratory for Acute Diarrheal Diseases of the Pedro Kourí Tropical Medicine Institute in Havana, Cuba. The timeline was subdivided into three five-year periods and one eight-year period. The centers submitting isolates were grouped into three geographical regions: western, central and eastern Cuba. A total of 1060 V. cholerae isolates were studied, from the 1438 samples sent from 15 Provincial Hygiene, Epidemiology and Microbiology Centers and the Municipal Hygiene, Epidemiology and Microbiology Center of the Isle of Youth Special Municipality. Genus, species and serotype of all specimens were studied and reviewed in the context of the outbreaks of acute diarrheal diseases reported in the country.
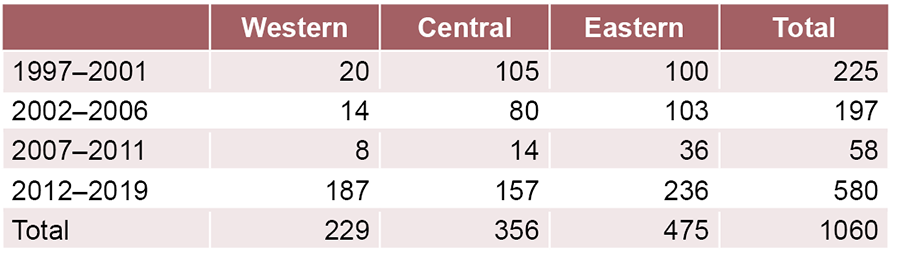
RESULTS All 1060 isolates were confirmed as V. cholerae. In the distribution by time period and region, the highest percentage occurred in the 2012‒2019 period, and the eastern region contributed the most isolates in all periods. Approximately 63.9% (677/1060) were from outbreaks, and in the 2012‒2019 period, the most epidemic-causing isolates came from the western region. Approximately 52.8% (560/1060) were identified as non-O1/non-O139 V. cholerae, and 47.2% (500/1060) as O1 V. cholerae; of these, 96.4% (482/500) corresponded to Ogawa serotype and 3.6% (18/500) to Inaba. Circulation of non-O1/non-O139 V. cholerae occurred throughout the entire period. The O1 serogroup began to circulate in 2012 and continued through 2016; however, since 2017, it has not been identified again. In the western region, there were smaller percentages of isolates of non-O1/non-O139 V. cholerae in all periods, except 2012‒2019. In that period, V. cholerae O1 was identified to a lesser degree in the central region.
CONCLUSIONSVibrio cholerae circulated in all three Cuban regions during the years studied, with a higher percentage of isolates of the non-O1/non-O139 serogroup, which caused outbreaks or sporadic cases of diarrhea in the eastern region, with the exception of the 2012‒2019 period, when epidemic outbreaks of the O1 serogroup (which causes cholera) occurred in all three regions, with higher percentages in the western region.
ABSTRACT INTRODUCTION Vibrio cholerae is a microorganism that causes acute diarrheal diseases and cholera, one of the leading causes of global morbidity and mortality, especially in children under five years old. It is present in many regions and has been isolated from diverse sources such as water, soil and food. Surveillance of this microorganism in Cuba from 1985 through June 1997 showed circulation of non-epidemic non-O1/non-O139 serogroups, but surveillance continued to identify distribution of V. cholerae serotypes and serogroups in the different geographic regions of the country during the following years, due to the risk of introducing cholera-causing serogroups that provoked cholera epidemics in other countries of the region.
OBJECTIVEDescribe the temporal‒spatial distribution of serogroups and serotypes of V. cholerae in Cuba.
METHODSA cross-sectional study was conducted that included isolates from passive surveillance of V. cholerae in 16 hygiene and epidemiology centers throughout Cuba from July 1997 through December 2019, submitted to the National Reference Laboratory for Acute Diarrheal Diseases of the Pedro Kourí Tropical Medicine Institute in Havana, Cuba. The timeline was subdivided into three five-year periods and one eight-year period. The centers submitting isolates were grouped into three geographical regions: western, central and eastern Cuba. A total of 1060 V. cholerae isolates were studied, from the 1438 samples sent from 15 Provincial Hygiene, Epidemiology and Microbiology Centers and the Municipal Hygiene, Epidemiology and Microbiology Center of the Isle of Youth Special Municipality. Genus, species and serotype of all specimens were studied and reviewed in the context of the outbreaks of acute diarrheal diseases reported in the country.
RESULTS All 1060 isolates were confirmed as V. cholerae. In the distribution by time period and region, the highest percentage occurred in the 2012‒2019 period, and the eastern region contributed the most isolates in all periods. Approximately 63.9% (677/1060) were from outbreaks, and in the 2012‒2019 period, the most epidemic-causing isolates came from the western region. Approximately 52.8% (560/1060) were identified as non-O1/non-O139 V. cholerae, and 47.2% (500/1060) as O1 V. cholerae; of these, 96.4% (482/500) corresponded to Ogawa serotype and 3.6% (18/500) to Inaba. Circulation of non-O1/non-O139 V. cholerae occurred throughout the entire period. The O1 serogroup began to circulate in 2012 and continued through 2016; however, since 2017, it has not been identified again. In the western region, there were smaller percentages of isolates of non-O1/non-O139 V. cholerae in all periods, except 2012‒2019. In that period, V. cholerae O1 was identified to a lesser degree in the central region.
CONCLUSIONSVibrio cholerae circulated in all three Cuban regions during the years studied, with a higher percentage of isolates of the non-O1/non-O139 serogroup, which caused outbreaks or sporadic cases of diarrhea in the eastern region, with the exception of the 2012‒2019 period, when epidemic outbreaks of the O1 serogroup (which causes cholera) occurred in all three regions, with higher percentages in the western region.
INTRODUCTION Acute diarrheal diseases—including cholera—are one of the main causes of global morbidity and mortality, especially in children under five years old.[1] Causal factors include various bacteria; among these, Vibrio cholerae is one of the most important.[2,3]
Morphologically, the microorganism has a curved structure with a polar flagellum.[4] Based on the O antigen of the capsular lipopolysaccharide, V. cholerae is classified in more than 200 biochemically identical serogroups, including O1 and O139, the two epidemic serogroups that cause cholera.[5] Non-O1/non-O139 serogroups are not considered epidemic, although they can cause outbreaks or sporadic cases of diarrhea with clinical symptoms and signs different from cholera.[3]
IMPORTANCEResults of microbiological surveillance for V. cholerae in Cuba provide useful and relevant information for the national health system and the Foodborne Diseases Program to channel the human resources and materials needed to control such acute diarrheal diseases.
V. cholerae is present in several regions of the world and can be isolated from diverse sources such as water, soil and food. Humans are an accidental and transitory host, yet they are the ones who disseminate it in the environment.[6,7] Intestinal infections are the most clinically relevant, particularly cholera,[8] but extra-intestinal infections can occur as well, such as fasciitis, bacteremia, meningitis, otitis and those that can develop in surgical wounds.[9]
Since 1991, in the region of the Americas, various cholera outbreaks have been reported[10] as well as sporadic cases of gastroenteritis or bacteremia caused by non-O1/non-O139 V. cholerae.[8,11,12] In 2010, acute diarrheal diseases caused by V. cholerae spread rapidly in Haiti,[13] a new scenario that could lead to reemergence and dissemination of cholera in countries such as the Dominican Republic, Cuba and Mexico, where positive cases were reported.[14,15] Considering circulation of this pathogen in the region, the National Reference Laboratory for Acute Diarrheal Diseases of the Pedro Kourí Tropical Medicine Institute (IPK) established in 1995 a surveillance system to identify outbreaks and sporadic cases of diarrheal illnesses that could be caused by V. cholerae.[16] This surveillance documented evidence of the circulation of non-O1/non-O139 strains from 1985 through June 1997, but surveillance of sporadic outbreaks of diarrheal diseases and epidemiological events possibly attributable to V. cholerae has continued, in order to provide relevant information to health authorities. The purpose of this study was to describe the temporal‒spatial distribution of V. cholerae serogroups and serotypes in Cuba from July 1997 through December 2019.
METHODS Design and samples A cross-sectional study was conducted from July 1997 through December 2019. It was subdivided into four periods: P1: 1997‒2001, P2: 2002‒2006, P3: 2007‒2011 and P4: 2012‒2019 (three five-year periods and one eight-year period, the latter in order to include the most recent results).
Of the 1483 V. cholerae isolates from passive surveillance in the culture collection of the IPK’s National Reference Laboratory for Acute Diarrheal Diseases, 1060 viable isolates (microorganisms with the possibility of multiplying) were studied; 423 resulted nonviable or were contaminated. The isolates were originally collected in 15 of Cuba’s Provincial Hygiene, Epidemiology and Microbiology Centers (CPHEM) and the Municipal Hygiene, Epidemiology and Microbiology Center (CMHEM) of the Isle of Youth Special Municipality.
The centers submitting isolates were grouped into three regions: western, central and eastern Cuba. The western region included CPHEMs of Pinar del Río, Artemisa, Mayabeque, Havana, and Matanzas provinces, as well as the CMHEM of the Isle of Youth Special Municipality; the central region included CPHEMs of Cienfuegos, Ciego de Ávila, Villa Clara, Sancti Spíritus and Camagüey provinces; and the eastern region included CPHEMs of Las Tunas, Holguín, Granma, Santiago de Cuba and Guantánamo provinces.
Isolates were classified according to genus, species, serogroup and serotype, and those related to outbreaks were identified. Cases were considered to be an outbreak when the unit submitting the V. cholerae isolates reported that they belonged to a single event in which two or more patients had been in contact with one another. The moment of onset of symptoms, the place where the episodes occurred, and the individuals’ characteristics were all recorded.[17]
Procedures Isolates were preserved in Pasteur’s conservation medium for Enterobacteriaceae; they were inoculated in brain-heart infusion broth and anaerobically incubated for 18‒24 hours at 37 °C. After incubation, a loopful of the broth culture was seeded by colony depletion plating on plates with selective medium for Vibrio thiosulfate-citrate-bile salts-sucrose (TCBS) agar (Biolife, Italy), MacConkey agar (Biolife, Italy) and blood agar (5% sheep blood) and incubated under recommended conditions.[18]
After incubating for 24 hours, three or more colonies were selected, based on their characteristics in the corresponding media: i) convex, with regular borders and sucrose fermenters in TCBS agar; ii) translucent, convex, with regular borders in MacConkey agar; and iii) hemolytic or not, convex, with regular borders in blood agar.
All colonies were inoculated by puncture and streaked in the primary differentiated media (Kligler’s iron agar with double sugar fermentation and lysine iron agar). Both were aerobically incubated from 18‒24 hours at 37 °C.[18]
Once incubation was complete, cultures that exhibited the following characteristics were selected: a) they did not oxidize or ferment lactose, they did oxidize and ferment glucose, they did not produce gas or hydrogen sulfide in Kligler’s iron agar iron with double sugar fermentation; and b) they decarboxylated L-lysine in iron and lysine agar.[18]
In all isolates, presence of cytochrome oxidase enzyme was confirmed, in accordance with the Kovacs method. Those that were oxidase positive were submitted to a complementary physiological study for confirmation in the Vibrio genus and V. cholerae species.[18]
To identify the genus, the Möeller method was utilized for biochemical tests measuring use of amino acids and carbohydrates.[18] To identify the species, isolates were submitted to tolerance tests for sodium chloride (NaCl) and use of sucrose. Serological agglutination tests were performed on slides using V. cholerae O1 and V. cholerae O139 polyvalent antisera to identify these serogroups.[18]
Analysis Descriptive statistics were used, such as frequency and percentages, to analyze and present results.
RESULTS The 1060 isolates were confirmed as V. cholerae as they were Gram-negative bacilli, facultative anaerobic, positive oxidase, with positive reaction to lysine and ornithine decarboxylase tests and negative for arginine dihydrolase; they grew in tryptone soy broth with 6% NaCl concentration and used sucrose and mannitol, not inositol.
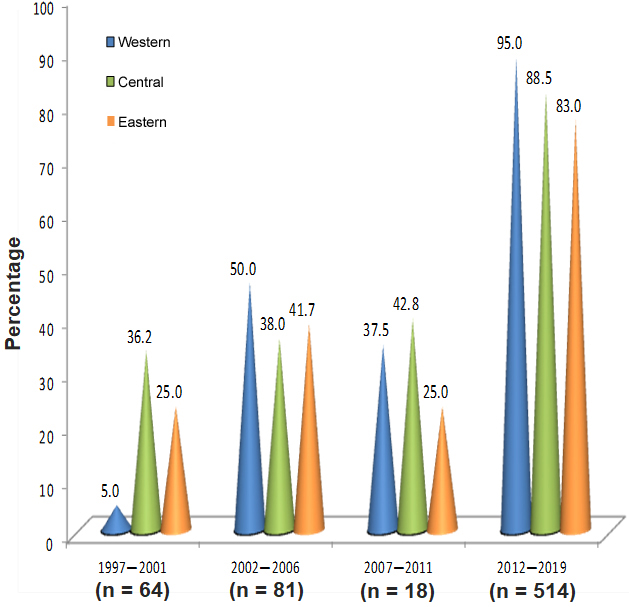
The highest percentage of V. cholerae isolates occurred in the 2012‒2019 period and in the eastern region in all periods (Table 1). Approximately 63.9% (677/1060) came from outbreaks. Figure 1 shows the proportion calculated for each period, according to geographic region. In the 1997‒2001 period, the lowest percentage of isolates coming from outbreaks was reported in the western region (5.0%; 34/677), but in the 2012‒2019 period, that region accounted for the highest proportion of isolates from outbreaks (95.0%; 643/677).
Of total isolates, 52.8% (560/1060) were identified as V. cholerae non-O1/non-O139 and 47.2% (500/1060) as V. cholerae O1, of which 96.4% (482/500) were Ogawa serotype and 3.6% (18/500) Inaba. None of the isolates were identified as V. cholerae O139.
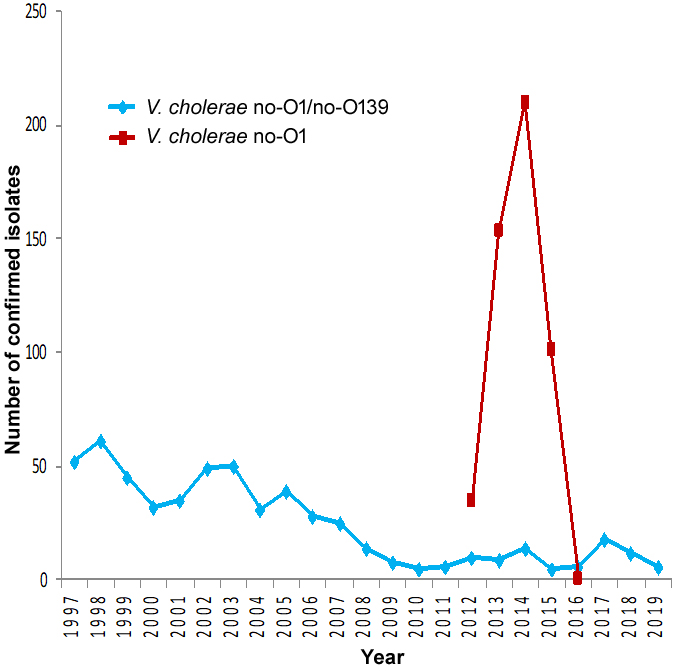
Figure 2 shows the temporal distribution of V. cholerae serogroups over the 23 years studied. As observed, V. cholerae non O1/non-O139 were in continuous circulation all those years. The O1 serogroup began circulating in 2012; the number of O1 isolates increased in 2013 and 2014, decreased in 2015, and since 2017, the O1 serogroup has not been reported at all.
Table 1: Distribution of confirmed isolates of Vibrio cholerae by geographic region. Cuba, July 1997‒December 2019
Figure 1: Proportion of isolates of Vibrio cholerae (n = 677) from outbreaks of diarrheal diseases by geographic region. Cuba, July 1997‒December 2019
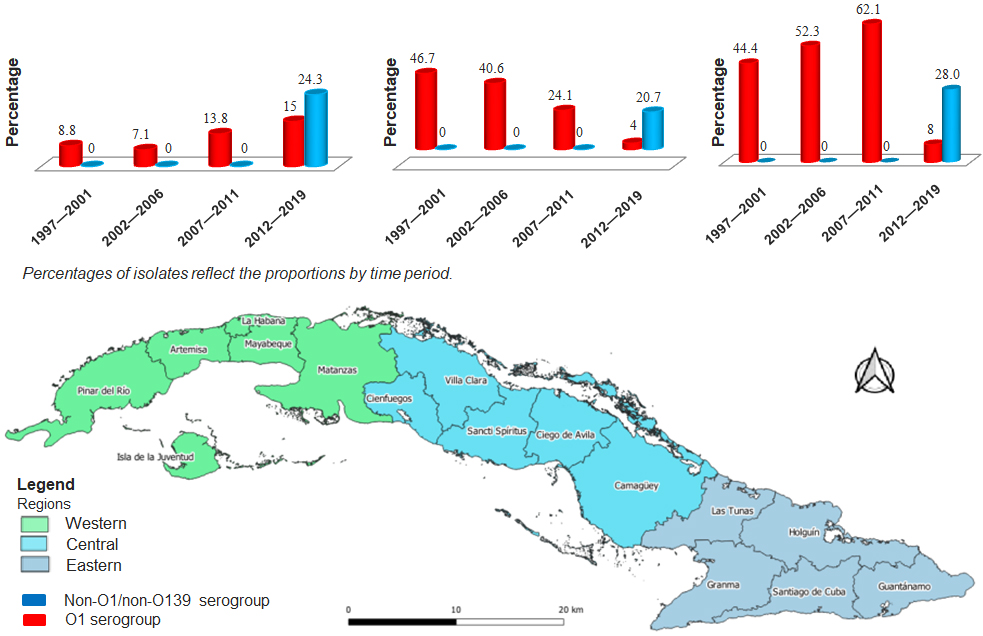
Figure 3 presents the distribution (in percentages) of serogroup strains according to geographic region. The western region presented the lowest percentage of non-O1/non-O139 V. cholerae isolates in all periods except 2012‒2019. In that same period, V. cholerae O1 was identified in the central region, but exhibiting the lowest percentages.
DISCUSSION Worldwide, Vibrio spp. are widely distributed in aquatic environments.[18] It has been suggested that as a result of climate change, increases in ocean temperatures may be responsible for outbreaks of Vibrio infections in countries such as Israel, Denmark, Spain, Chile and the United States.[19,20]
According to data published by the Global Burden of Diarrheal Diseases Collaborators (GBDDC), acute diarrheal diseases have persisted in Latin America as a significant public health problem.
Figure 2: Distribution over time of Vibrio cholerae O1 serogroup and Vibrio cholerae non-O1/non-O139 serogroup (n = 1060) responsible for diarrheal diseases. Cuba, July 1997‒December 2019
[21] Levels during the last three decades have been relatively stable and mortality in several countries has dropped, which Herrera-Benavente attributes to the success of WHO-sponsored control programs (under auspices of PAHO).[1]
In a study prior to implementation of Cuba’s surveillance system, Bravo[16] found that in the three Cuban regions, non-O1/non-O139 V. cholerae circulated from 1985 through 1997 in patients with acute diarrheal diseases, most commonly in the eastern region. The author suggested that an ecological niche in that region might favor persistence of diarrheas caused by this agent.
In June 2012, V. cholerae O1 was isolated in Cuba, when the Ministry of Public Health reported a cholera outbreak in Granma province.[22] Although no events caused by this strain had been observed in previous years, it is not unusual for such events to occur. The cholera-causing agent cannot be eliminated from the environment since it is a native species and is associated with marine habitats such as algae and crustaceans. In addition, changes in climate patterns favor the growth of V. cholerae in aquatic ecosystems and contribute to the occurrence of outbreaks and epidemics.[23,24]
In Cuba, more outbreaks and cases of diarrhea occur in the summer months, in part due to high temperatures and summer rains.[25] However, no studies have been conducted on Cuban climate variability and its effect on the temporal‒spatial behavior of V. cholerae.
The epidemic O1 and O139 V. cholerae serogroups produce cholera, particularly in developing countries due to faulty infrastructure affecting basic sanitation and drinking water, as well as poor access to quality medical services,[2,26,27] while non-O1/non-O139 strains are often isolated in the environment and are associated with sporadic cases of gastroenteritis and extraintestinal infecctions[28]
Figure 3: Distribution of Vibrio cholerae serogroup O1 and Vibrio cholerae non-O1/non-O139 responsible for diarrheal diseases (n = 1060) according to geographic region.
Cuba, July 1997‒December 2019
From 2012 to 2016, there were various outbreaks in Cuba associated with the circulation of V. cholerae O1.[22,29] In the present study, an increase in isolates was observed from 2012 to 2019 from outbreaks in the country’s western region, which could be related to the joint circulation of the epidemic serotypes that can cause outbreaks of greater magnitude.[27]
In a 2012 study in Cuba by Romero-Placeres to investigate whether shortcomings in the quality of drinking water and sanitation influenced the burden of acute diarrheal diseases, it was found that more municipalities in the west faced high risk of a diarrheal outbreak due to both these problems. In addition, Havana is located in western Cuba, and is the province with the largest population and most dealings (through business, tourism, travel) with the rest of the island and also with other countries.[30] Another important factor is that 96.8% of households in the capital receive water via aqueduct; the supply service is inconsistent and in some places has low pressure, which along with the current poor condition of distribution pipes, can lead to water contamination.[31]
According to the 2012 census by Cuba’s National Statistics Bureau,[31] despite the fact that the country enjoys extensive coverage of water and sanitation services, it still faces deficiencies from deteriorating infrastructure due to aging distribution systems, with high risk of microbiological contamination, a conclusion coinciding with observations by Romero-Placeres.[30]
The 2012 and 2013 cholera outbreaks in Havana were associated with food markets, where several asymptomatic carriers worked and manipulated foods.[25] The O1 serogroup contains three serotypes; the two most common—Ogawa and Inaba—are capable of producing cholera outbreaks. The difference between the two lies in methylation of the capsular lipopolysaccharide by the enzyme methyltransferase, codified by the webT gene. When an isolate presents a methylated lipopolysaccharide, it belongs to the Ogawa serotype, and when it does not, it is Inaba.[32]
In endemic areas, the change from one serotype to another is not a random process, rather, it depends on several factors.[33] Genetic studies suggest that the mobile elements present in the genome of V. cholerae, such as those related to virulence (CTXØ), antibiotic resistance (SXT ICE) and phages and their resulting clonal expansion, can contribute to a preponderance of a particular serotype during a specific time period. Another important factor in the change of serotype is pressure generated by the immune system, because when a specific serotype circulates in a population over a long time, the host will present immunity to this serotype. The effect of environmental factors on the viability of these two serotypes is not well known.[34]
In stool samples of patients with diarrhea obtained from 2006 through 2016 in sentinel centers in Nepal, V. cholerae O1 was isolated, mostly the Ogawa serotype,[35] and in 2015 and 2016 this strain was also isolated in stool samples of patients with acute diarrheal disease and identified in samples of water used for drinking and domestic chores in Ghana, coexisting with non-O1/non-O139 serogroup.[36]
During cholera outbreaks in Cuba, circulating together with V. cholerae O1 were the non-epidemic serogroups, as reported in Thailand, Iraq and Japan.[37] The coexistence of toxigenic and nontoxigenic serogroups represents a global health emergency, due to the horizontal transfer of genes that contribute virulence and antimicrobial resistance among serogroups of the same species, which can lead to the appearance of more virulent strains.[38]
The results of the present study on circulation of serogroups and serotypes coincided with the above-cited works and with cocirculation of V. cholerae O1 and non-O1/non-O139 from 2012 to 2016, underscoring the importance of epidemiological surveillance of this bacterium in Cuba.
In January 1991, a cholera epidemic broke out on the coast of Peru, attributed to the O1 serogroup, which quickly spread to almost all Latin American countries, although Cuba was not affected.[25] In October 2010, cholera was introduced into Haiti, and from then through 2018, persistent circulation and outbreaks of cholera were reported, associated with poor sanitary conditions and lack of access to safe drinking water in that country.[13] Subsequent outbreaks in Cuba were related to the Haitian epidemic, according to Cuban health officials, who cited the ongoing interaction between the two countries.[39]
During the period studied, V. cholerae O139 was not reported in Cuba. This serogroup, first identified in 1992 in Bangladesh, has caused outbreaks elsewhere in the past, but in recent years it has been reported only in sporadic cases, always in the Asian region.[27]
The principal limitation of this study is that unidentified outbreaks may have occurred in the country’s rural zones and cases not included because the infected individuals did not seek treatment in a medical center.
CONCLUSIONS Our temporal‒spatial heterogeneity study confirmed that V. cholerae circulated for 23 years in all three regions of Cuba. Additionally, non-O1/non-O139 serogroups that caused outbreaks or sporadic cases of diarrhea were isolated in all time periods and regions, most often in the eastern region, with the exception of the last period, when epidemic outbreaks of the cholera-causing O1 strain occurred in all three regions, most commonly in the western region. This study shows that gastrointestinal infections from V. cholerae constitute a health problem in Cuba that needs to be addressed and suggests that the results should be used to design focused strategies, in accordance with the microorganism’s temporal-spatial distribution pattern.
Herrera-Benavente IF, Comas-García A, Mascareñas-de los Santos AH. Impacto de las enfermedades diarreicas agudas en América Latina. Justificación del establecimiento de un Comité de Enfermedades Diarreicas en SLIPE. Rev Latin Infect Ped. 2018 Jan–Mar;31(1):8–16. Spanish.
Parte AC. LPSN—list of prokaryotic names with standing in nomenclature (bacterio.net), 20 years on. Int J Syst Evol Microbiol. 2018 Jun 1;68(6):1825–9. https://doi.org/10.1099/ijsem.0.002786
Farmer JJ, Janda JM, Brenner FW, Cameron DN, Birkhead KM. Vibrio. In: Bergey’s Manual of Systematics of Archaea and Bacteria. New Jersey: John Wiley & Sons, Inc., Bergey’s Manual Trust; 2015 Apr 17. https://doi.org/10.1002/9781118960608.gbm01078
Shin OS, Tam VC, Suzuki M, Ritchie JM, Bronson RT, Waldor MK, et al. Type III secretion is essential for the rapidly fatal diarrheal disease caused by non-O1, non-O139 Vibrio cholerae. mBio. 2011 Jun 14;(3)2:e00106-11. https://doi.org/10.1128/mbio.00106-11
Chin J, editor. El control de las enfermedades transmisibles. 17 ed [Internet]. Washington, D.C.: Pan American Health Organization; 2001 [cited 2018 Jan 5]. 748 p. Available at: https://www.paho.org/hq/dmdocuments/2010/9275315817.pdf. Spanish.
Olivares F, Domínguez I, Dabanch J, Porte L, Ulloa MT, Osorio G. Bacteriemia por Vibrio cholerae no-O1/no-O139 que porta una región homóloga a la isla de patogenicidad VpaI-7. Rev Chil Infectol. 2019 Jun;36(3):392–5. https://doi.org/10.4067/S0716-10182019000300392. Spanish.
Hirk S, Huhulescu S, Allerberger F, Lepuschitz S, Rehak S, Weil S, et al. Necrotizing fasciitis due to Vibrio cholerae non-O1/non-O139 after exposure to Austrian bathing sites. Wien Klin Wochenschr. 2016 Feb 29;128(3–4):141–5. https://doi.org/10.1007/s00508-015-0944-y
Domman D, Quilici ML, Dorman MJ, Njamkepo E, Mutreja A, Mather AE, et al. Integrated view of Vibrio cholerae in the Americas. Science. 2017 Nov 10;358(6364):789–93. https://doi.org/10.1126/science.aao2136
Montero D, Vidal M, Pardo M, Torres A, Kruger E, Farfán M, et al. Characterization of enterotoxigenic Escherichia coli strains isolated from the massive multi-pathogen gastroenteritis outbreak in the Antofagasta region following the Chilean earthquake, 2010. Infect Genet Evol. 2017 Aug;52:26–9. https://doi.org/10.1016/j.meegid.2017.04.021
Arteaga M, Velasco J, Rodríguez S, Vidal M, Arellano C, Silva F, et al. Genomic characterization of the non-O1/non-O139 Vibrio cholerae strain that caused a gastroenteritis outbreak in Santiago, Chile, 2018. Microb Genom. 2020 Mar;12:3. https://doi.org/10.1099/mgen.0.000340
Guillaume Y, Raymond M, Jerome GJ, Ternier R, Ivers LC. ‘It was a ravage!’: lived experiences of epidemic cholera in rural Haiti. BMJ Glob Health. 2019 Nov 10;16(4):e001834. https://doi.org/10.1136/bmjgh-2019-001834
Estrada-Paneque MU, Estrada-Vinajera G, Vinajera-Torres C. Cólera. Contexto clínico-epidemiológico y social. A propósito del brote epidémico en Granma, Cuba. Multimed [Internet]. 2017 [cited 2019 Mar 10]. Available at: http://www.revmultimed.sld.cu/index.php/mtm/article/view/601. Spanish.
Pan American Health Organization, World Health Organization. Actualización Epidemiológica: Cólera 23 de nov 2016. Washington, D.C.: Pan American Health Organization, World Health Organization; 2016. Spanish.
Bravo L. Estudio del género Vibrio en Cuba [doctoral thesis]. [Havana]: Pedro Kourí Institute of Tropical Medicine (CU); 2002. Spanish.
Peláez O, Más P. Brotes, epidemias, eventos y otros términos epidemiológicos de uso cotidiano. Rev Cubana Salud Pública. 2020;46(2):23–35. Spanish.
Koneman E, Win W, Allen S, Janda W, Procop G, Schreckenberg P. Koneman Diagnóstico microbiológico. Texto y Atlas en color. 6th ed. Madrid: Editorial Médica Panamericana; 2007. p. 129, 190, 372. Spanish.
Baker-Austin C, Trinanes J, González-Escalona N, Martínez-Urtaza J. Non-cholera vibrios: the microbial barometer of climate change. Trends Microbiol. 2017 Jan 1;25(1):76–84. https://doi.org/10.1016/j.tim.2016.09.008
Di DYW, Lee A, Jang J, Han D, Hur HG. Season-specific occurrence of potentially pathogenic Vibrio spp. on the southern coast of South Korea. Appl Environ Microbiol. 2017 Jan 17;83(3):e02680-16. https://doi.org/10.1128/AEM.02680-16
Global Burden of Diarrhoeal Diseases Collaborators (GDDC). Estimates of global, regional, national morbidity, mortality, and aetiologies of diarrhoeal diseases: a systematic analysis for the Global Burden of Disease Study 2015. Lancet Infect Dis. 2017 Sep;17(9):909–48. https://doi.org/10.1016/s1473-3099(17)30276-1
Chowdhury FR, Nur Z, Hassan N, von Seidlein L, Dunachie S. Pandemics, pathogenicity and changing molecular epidemiology of cholera in the era of global warming. Ann Clin Microbiol Antimicrob. 2017 Mar 7;16:10. https://doi.org/10.1186/s12941-017-0185-1
Froelich BA, Daines DA. In hot water: effects of climate change on Vibrio-human interactions. Environ Microbiol. 2020 Oct;22(10):4101–11. https://doi:10.1111/1462-2920.14967
Battle MC, Dickinson F. Notas para una historia del cólera en Cuba durante los siglos XIX, XX, XXI. Rev Anales Acad Cien Cuba. 2014;4(1):1–11. Spanish.
Jameel SK, Shafek MA, Abdulmohsen AM, Mohamed N, Sarmad RN, Tuhama TM. The isolation of Vibrio cholerae and other enteric bacteria with molecular characterization of V. cholerae during outbreak of Baghdad/Iraq in 2015. Adv Microb. 2016 Aug;6(9):699–715. http://dx.doi.org/10.4236/aim.2016.69069
World Health Organization [Internet]. Geneva: World Health Organization; c2022. Centro de Prensa. Cólera. Datos y cifras; 2021 Aug 17 [cited 2021 Dec 24]; [about 3 p.]. Available at: https://www.who.int/es/news-room/fact-sheets/detail/cholera/. Spanish.
Ulloa MT, Sanhueza C, Henríquez T, Aguayo B, Hermosilla G, Porte L, et al. [Chilean strains of clinical origin of non-O1, non-O139 Vibrio cholerae carry the genes vcsN2, vcsC2, vcsV2, vspD, toxR2 y vopF from secretion system T3SS2 present in an island of pathogenicity]. Rev Chilena Infectol. 2019 Jun;36(3):312–7. https://doi.org/10.4067/s0716-10182019000300312. Spanish.
Zelada-Valdés A, Ledón-Pérez TY, Fando-Calzada RA. El cólera: una enfermedad infecciosa reemergente. El candidato vacunal CV 638, una herramienta para su prevención. Rev CENIC. 2015 May–Aug;46(2):131–43. Spanish.
Romero-Placeres M. Estimación de la carga de enfermedad para enfermedades diarreicas relacionadas al agua y saneamiento en municipios seleccionados de Cuba en 2012 [doctoral thesis]. [México, DF]: National Institute of Public Health (MX), National School of Public Health of Mexico; 2015. Spanish.
Liang W, Wang L, Liang P, Zheng X, Zhou H, Zhang J, et al. Sequence polymorphisms of rfbT among the Vibrio cholerae O1 strains in the Ogawa and Inaba serotype shifts. BMC Microbiol. 2013 Jul 26;13:173. https://doi.org/10.1186/1471-2180-13-173
Karlsson SL, Thomson N, Mutreja A, Connor T, Sur D, Ali M, et al. Retrospective analysis of serotype switching of Vibrio cholerae O1 in a cholera endemic region shows it is a non-random process. PLoS Negl Trop Dis. 2016 Oct 5;10(10):e0005044. https://doi.org/10.1371/journal.pntd.0005044
Baddam R, Sarker N, Ahmed D, Mazumder R, Abdullah A, Morshed R, et al. Genome dynamics of Vibrio cholerae isolates linked to seasonal outbreaks of cholera in Dhaka, Bangladesh. mBio. 2020;11(1):e03339-19. https://doi.org/10.1128/mBio.03339-19
Rijal N, Acharya J, Adhikari S, Upadhaya BP, Shakya G, Kansakar P, et al. Changing epidemiology and antimicrobial resistance in Vibrio cholerae: AMR surveillance findings (2006-2016) from Nepal. BMC Infect Dis. 2019 Sep 11;19(1):801. https://doi.org/10.1186/s12879-019-4432-2
Abana D, Gyamfi E, Dogbe M, Opoku G, Opare D, Boateng G, et al. Investigating the virulence genes and antibiotic susceptibility patterns of Vibrio cholerae O1 in environmental and clinical isolates in Accra, Ghana. BMC Infect Dis. 2019 Jan 21;19(1):76. https://doi.org/10.1186/s12879-019-3714-z
Wang H, Yang C, Sun Z, Zheng W, Zhang W, Yu H, et al. Genomic epidemiology of Vibrio cholerae reveals the regional and global spread of two epidemic non-toxigenic lineages. PLoS Negl Trop Dis. 2020 Feb 18;14(2):e0008046. https://doi.org/10.1371/journal.pntd.0008046
Valáriková J, Korcová J, Ziburová J, Rosinský J, Čížová A, Bieliková S, et al. Potential pathogenicity and antibiotic resistance of aquatic Vibrio isolates from freshwater in Slovakia. Folia Microbiol. 2019 Nov 26. https://doi.org/10.1007/s12223-019-00760-w
Mena VR, Ávila I, Cabrera AC. Caracterización de las complicaciones de los pacientes diagnosticados con cólera en el Hospital Pediátrico de Centro Habana, 2013-2017. Rev Cubana Pediatr. 2020 Jan–Mar;92(1):e919. Spanish.
THE AUTHORS
Anabel Fernández-Abreu (Corresponding author: anabel@ipk.sld.cu), chemical engineer with a master’s degree in bacteriology and a doctorate in health sciences. Associate researcher, National Reference Laboratory for Acute Diarrheal Diseases, Pedro Kourí Tropical Medicine Institute (IPK), Havana, Cuba. https://orcid.org/0000-0002-5395-5041
Laura Bravo-Fariñas, biologist with a doctorate in health sciences. Consulting professor and senior researcher, IPK, Havana, Cuba. https://orcid.org/0000-0003-2183-3119
Yanaika Cruz-Infante, health technologist with a master’s degree in bacteriology-mycology. Adjunct researcher, IPK, Havana, Cuba. https://orcid.org/0000-0002-9825-2737
Rosabel Falcón-Márquez, microbiologist with a doctorate in health sciences. Senior researcher, IPK, Havana, Cuba. https://orcid.org/0000-0003-1273-6835
María Eugenia Toledo-Romaní, epidemiologist with a doctorate in medical sciences. Senior researcher, IPK, Havana, Cuba. https://orcid.org/0000-0001-8600-9062
María de los Ángeles León-Venero, epidemiologist. Head, Foodborne Diseases Program, National Epidemiology Division, Ministry of Public Health, Havana, Cuba. https://orcid.org/0000-0002-2962-4090
No new Early Online articles at this time. The most recent articles are listed in the Current Issue Table of Contents. Early Online articles are added as soon as they are available, so please check back later.