INTRODUCTION Most previous studies have examined the effects of acute psychological stress in humans based on select gene panels. The genomic approach may help identify novel genes that underline biological mechanisms of acute psychological stress responses.
OBJECTIVE This exploratory study aimed to investigate genome-wide transcriptional activity changes in response to acute psychological stress.
METHODS The sample included 40 healthy women (mean age 31.4 ± 11.6 years). Twenty-two participants had a stress experience induced by the Trier Social Stress Test (experimental group) and 18 did not (control group). Psychological stress levels and hemodynamic changes were assessed before and after the Trier Social Stress Test. Peripheral blood samples obtained before and after the Trier Social Stress Test were processed for mRNA sequencing.
RESULTS Psychological and hemodynamic stress parameters indicated that the Trier Social Stress Test induced moderate evels of stress in the experimental group. Six genes (HCG26, HCP5, HLA-F, HLA-F-AS1, LOC1019287, and SLC22A16) were up-regulated, and five genes (CA1, FBXO9, SNCA, STRADB, and TRMT12) were down-regulated among those who experienced stress induction, compared with the control group. Nine genes of eleven were linked to endocrine system disorders, neurological disease, and organismal injury and abnormalities.
CONCLUSION Of the genes identified in this study, HCP5, SLC22A16, and SNCA genes have previously been proposed as therapeutic targets for cancer and Parkinson disease. Further studies are needed to examine pathological mechanisms through which these genes mediate effects of psychological stress on adverse health outcomes. Such studies may ultimately identify therapeutic targets that enhance biological resilience to adverse effects of psychological stress.
KEYWORDS Stress, psychological; sequence analysis, RNA; psychological tests; US
ABSTRACT INTRODUCTION Most previous studies have examined the effects of acute psychological stress in humans based on select gene panels. The genomic approach may help identify novel genes that underline biological mechanisms of acute psychological stress responses.
OBJECTIVE This exploratory study aimed to investigate genome-wide transcriptional activity changes in response to acute psychological stress.
METHODS The sample included 40 healthy women (mean age 31.4 ± 11.6 years). Twenty-two participants had a stress experience induced by the Trier Social Stress Test (experimental group) and 18 did not (control group). Psychological stress levels and hemodynamic changes were assessed before and after the Trier Social Stress Test. Peripheral blood samples obtained before and after the Trier Social Stress Test were processed for mRNA sequencing.
RESULTS Psychological and hemodynamic stress parameters indicated that the Trier Social Stress Test induced moderate levels of stress in the experimental group. Six genes (HCG26, HCP5, HLA-F, HLA-F-AS1, LOC1019287, and SLC22A16) were up-regulated, and five genes (CA1, FBXO9, SNCA, STRADB, and TRMT12) were down-regulated among those who experienced stress induction, compared with the control group. Nine genes of eleven were linked to endocrine system disorders, neurological disease, and organismal injury and abnormalities.
CONCLUSION Of the genes identified in this study, HCP5, SLC22A16, and SNCA genes have previously been proposed as therapeutic targets for cancer and Parkinson disease. Further studies are needed to examine pathological mechanisms through which these genes mediate effects of psychological stress on adverse health outcomes. Such studies may ultimately identify therapeutic targets that enhance biological resilience to adverse effects of psychological stress.
KEYWORDS Stress, psychological; sequence analysis, RNA; psychological tests; US
INTRODUCTION Psychological stress is associated with adverse health outcomes, particularly with depression, cardiovascular disease and cancer.[1] While many studies have demonstrated a potential link between chronic stress and altered inflammatory cytokine levels, immune dysfunctions and delayed neuronal recovery,[2,3] it is still unclear how acute stress is processed at transcriptional levels. Recently, a study conducted in mice demonstrated that a single stress event left long-lasting changes in microRNA, messenger RNA and protein expressions in the amygdala.[4] Although many animal studies have attempted to determine the mechanisms underlying stress pathophysiology, human studies investigating subcellular responses to acute psychological stress are limited. In addition, it is recognized that gene expression in animal models is not readily translated to humans.[5]
Available data collected in humans demonstrate that acute psychological stress is a potent trigger of inflammatory, neuroendocrine and metabolic responses, resulting in predisposition to disease. For example, acute stress increases proinflammatory gene expres sion, including interleukin 6 (IL-6), interleukin 1 beta (IL-1β), type 1 T-helper/type 2 T-helper (TH1/TH2) cytokines, catecholamine receptors, nuclear factor kappa beta (NF-kB), I kappa beta kinase (IкB), or tumor necrosis factor alpha (TNF-α);[6-10] the hypothalamic–pituitary–adrenal (HPA) axis-related genes;[11] immune activity;[12,13] and glucose metabolism.[14] However, most studies have investigated expression of only a few select genes, and only a limited number of studies have examined genome-wide transcriptional activity in response to acute psychological stress using microarrays.[15,16] The RNA-sequencing (RNA-seq) genomic approach to identifying differentially-expressed genes may contribute to a better understanding of the biological mechanisms underlying acute psychological stress responses. It may also identify novel genes that influence previously known and unknown stress response pathways. We investigated the effect of acute psychological stress on genome-wide transcriptome profiles in whole-blood samples via RNA-seq and explored which genes may be differentially expressed after stress induction using a randomized controlled design. We used the Trier Social Stress Test (TSST), a standard laboratory procedure used to reliably induce stress in human research participants.[17,18] This exploratory approach may identify genes that can then be studied in future confirmatory work.
IMPORTANCE This study highlights the need to understand the role of HCP5, SLC22A16, and SNCA genes in linking psychological stress to cancer and Parkinson disease.
METHODS Participants All measurements and blood samples were obtained during a previous study, which evaluated changes in arterial stiffness after stress induction.[19] After institutional review board approval for the previous study, a convenience sample of 85 women was recruited from Charlottesville, Virginia, USA, communities, using flyers, emails and word of mouth. The study included female adults aged 18–55 years. This selected sex and age range reduced sample heterogeneity and obviated the need to control for potential confounding effects of older age and menopause on cardiovascular function. To minimize potential confounding effects, women were also excluded who were taking medications for psychological issues (e.g., anxiolytics and antipsychotics), had any diagnosed cardiovascular disease (e.g., hypertension, diabetes mellitus or hyperlipidemia), or had gone through menopause. In the previous study, participants were randomized by coin flips into either the intervention (stress induction) or control (no stress induction) group. Financial considerations prohibited genetic sequencing on all 85 participants. Because the previous study aimed to test changes in transcriptome profiles in relation to arterial stiffness before and after stress induction, only the 40 participants who had exhibited changes in arterial stiffness were selected for genetic sequencing; 22 received the experiment (experimental group), and 18 did not (control group). This sample was used in the current study to analyze transcriptome changes related to acute psychological stress induction.
Ethics All procedures were conducted after the approval from the Institutional Review Board at University of Virginia, USA. Written informed consent was obtained from all patients included in the study.
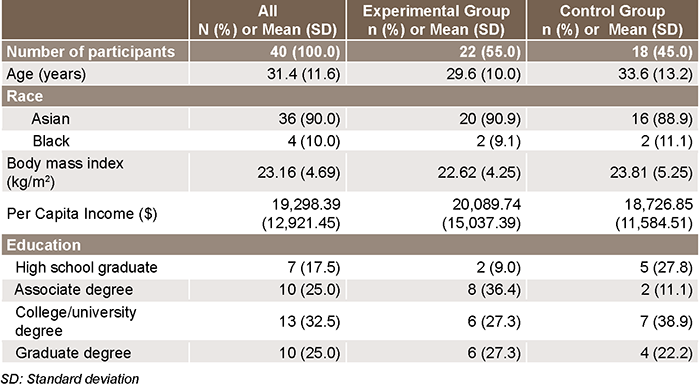
Background Age, race, per capita income and education level were self-reported by participants.
Psychological factors Subjective acute psychological stress levels were measured by the Subjective Units of Distress Scale (SUDS)[20] and the state anxiety subscale of the Spielberger State-Trait Anxiety Inventory (STAI).[21]
Subjective Units of Distress Scale: Current and peak distress was measured by SUDS, widely-used one-item scale.[20] The question asked before the experiment (for the experimental group) or the sitting period (for the control group) was: “On a scale of 0 to 10, with 0 ‘not distressed at all’ and 10 ‘the most distressed’, what is your distress level now?” The question asked after the experiment (or after the sitting period) was: “On a scale of 0 to 10, with 0 ‘not distressed at all’ and 10 ‘the most distressed’, what was your highest level of distress during the experiment or the sitting period?”.
State anxiety subscale of the Spielberger State-Trait Anxiety Inventory: State anxiety was also measured before and after the experiment (or after the sitting period for the control group). The 20-item state anxiety subscale uses a 4-point Likert-like scale to assess anxiety intensity, tied to “how one feels right now, that is, at this moment” (e.g., feelings of worry, tension). The total score is derived from the sum of the items with higher scores indicating greater anxiety.[21] This scale has been used extensively and has strong construct and divergent validity and internal consistency in large samples.[22] In the current study, baseline state anxiety scale internal consistency was excellent (Cronbach’s α = 0.90).
Physiological factors Mean arterial pressure and heart rate Blood pressure (BP) and heart rate (HR) are commonly used to assess physiological responses to stress. In this study, BP and HR were included as indicators of acute psychological stress, along with SUDS and STAI’s state anxiety subscale. BP and HR were measured using the Welch Allyn Vital Signs Monitor 300 Series (Welch Allyn, USA). After measuring mid-section circumference of the nondominant upper arm, the proper-sized cuff was applied snuggly with the artery marker on the cuff placed over the brachial artery.[23] BP was measured on the arm kept still, at the level of the heart. Given that concurrent use of systolic BP and diastolic BP may cause multicollinearity issues, mean arterial blood pressure (MAP) was used by calculating diastolic BP +1/3 (systolic BP–diastolic BP).
Body mass index (BMI) This is an anthropometric measurement that may affect physiological response to stress;[24] thus, BMI was included as a covariate in transcriptome analysis. Height (m) and weight (kg) were measured to determine BMI (kg/m2). Height was measured using a wall stadimeter (Accu-Hite, USA), and weight was measured with an electronic scale (Penn Scale, USA).
Procedures Participants’ physiological conditions were standardized according to the European Society of Cardiology’s recommendations for measuring cardiovascular function.[25,26] To minimize variance caused by circadian patterns, study procedures were conducted in a quiet room between 1:00 PM and 3:00 PM. All participants refrained from vigorous exercise and from consuming coffee, tea, bananas, chocolate, cocoa, citrus fruits and vanilla for one day before data collection, because vigorous exercise and consumption of such foods may change cardiovascular hemodynamics. Participants ate the same breakfast of cereal (35 g), milk (250 mL) and orange juice (250 mL) at 8:00 AM, followed by a fasting period until data collection was completed in the afternoon.
Pre-test data collection For baseline stress measurements, participants completed the SUDS and the STAI subscale. Next, height and weight were measured. After 10 minutes resting in the supine position, BP was measured. Blood was drawn by an experienced nurse–phlebotomist. Participants were then randomized by coin flip to either experimental or control group.
Trier Social Stress Test (TSST) For the experimental group, acute psychological stress was induced by using the TSST, which is a valid and reliable tool used to induce acute psychological stress and to study biological responses to stress in laboratory settings.[17,18] The intention of the TSST is to create perceived uncontrollability and fears of negative social evaluation. These two components are considered central in biological stress reactivity activation, such as activation of the hypothalamic-pituitary-adrenal (HPA) axis and sympathetic adrenal–medullary axis.[27] The TSST requires speech performance and verbal arithmetic performance in front of an audience, and is known to induce considerable changes in corticotropin,[28] cortisol,[29,30] blood pressure [29] and heart rate.[31]
The TSST was conducted in an intervention room located across the hall from the room where baseline measures were collected. Individuals assigned to the experimental group were given instructions for the upcoming TSST task. They had 10 minutes to prepare a 5-minute speech in which they were asked to convince two interviewers that they were a strong candidate for their dream job. Participants were told that their performances would be videotaped and evaluated by interviewers. Participants stood in front of a video camera and two interviewers (research staff dressed in lab coats). During the presentation, the interviewers maintained neutral expressions and periodically took notes. If participants stopped early, they were encouraged to continue. If a participant repeatedly looked at the interviewers or stopped talking for more than 10 seconds, they were told that “you have X minutes left but you can let me know if you wish to stop”. Each presentation was immediately followed by a mental arithmetic test, which involved sequentially subtracting the number 13 from 1022. If a mistake was made, they were asked to start again from the beginning. This continued until the participant had completed the task for the full five minutes.[17] Participants in the control group spent about 20 minutes quietly sitting in the same intervention room without undergoing the TSST.
Post-test data collection Immediately after the TSST period or the sitting period, participants in both groups completed the SUDS a second time. Next, in a supine position, BP was measured, and peripheral blood was drawn again from peripheral catheters, which had been placed earlier. Finally, participants were asked to complete the STAI subscale again before being fully debriefed and compensated for their participation.
RNA extraction, library preparation and sequencing Peripheral blood (2.5 mL) was collected in a PAXgene RNA tube (QIAGEN, USA) and stored at –80 ˚C until ready for RNA extraction. RNA was extracted from whole blood using the PAXgene Blood RNA system (QIAGEN, USA). RNA sample quality was evaluated using an Agilent 4200 TapeStation (Agilent Technologies, USA) by the RNA Integrity Number (RIN), and the quantity of RNA was measured using a Qubit (Life Technologies, USA). All samples used for this study had excellent purity (A260/A280 ≥1.9; A260/A230 ≥2) and showed no visible signs of degradation (RIN ≥9). We used the TruSeq Stranded mRNA library prep kit (Illumina, USA) to generate mRNA-sequencing libraries. These kits generated high-quality libraries for sequencing by fragmentizing RNA, performing reverse transcription and ligating the indexed adapters. This allowed individual libraries to be pooled in an equimolar fashion, minimizing the potential technical bias of run variation. Pooled libraries were then sequenced with an Illumina NextSeq 500 instrument (Illumina, USA).
Analysis Study participant characteristics are described by means and standard deviation (SD) for continuous variables, and by frequency and percent for categorical variables. Condition differences in participants’ characteristics at baseline were examined using independent sample t-tests. To compare psychological and physiological responses between the experimental and control groups, repeated measures of covariance analyses (RM-ANCOVAs) were conducted; controlling for factors which may affect physiological responses to stress (age, MAP and BMI). The within-subjects factor was ‘time’ (pre- and post-stressor), and the between-subjects factor was ‘stressor’ (TSST and control). Interactions of within-subjects factor and between-subjects factor were tested. All statistical analyses were performed with SPSS Statistics 25 for Windows (SPSS, USA).
Bioinformatic analysis of RNA-sequencing data We performed bioinformatics quality control using FastQC, version 0.11.7 (Babraham Bioinformatics, UK). Poor quality reads and adapter sequences were filtered out by running CutAdapt, version 2.5.[32] To confirm the quality of the library and sequencing, we used RNA-SeQC[33] for quality control specific to RNA-sequencing, and assessed total number of reads, depth of reads, average read length, average coverage across the gene, number of identified genes, PCR duplication rate, ribosomal content and exon/intron representation. We aligned the raw reads to the GRCh38 reference genome using STAR version 2.6.1a.[34] We counted number of reads mapped to genes using HTSeq, version 0.11.0.[35] We performed differential gene expression analyses between experimental and control groups using DESeq2, version 1.30.1 [36] while controlling for age, MAP and BMI as potential covariates. The gene count table was imported to DESeq2. Read distribution was modeled as a negative binomial distribution with mean and variance estimated from data. P values were calculated by using the Wald test. Multiple testing correction was performed with Benjamini-Hochberg’s False Discovery Rate (FDR) adjusted by the Independent Hypothesis Weighting method with a cutoff of 0.05 (on FDR),[37] accounting for age, BMI and MAP as covariate variables. R version 4.0.3 (2020-10-10) and BiocManager version 3.12 (Bioconductor, USA) were used.
Construction of pathway–gene–process network Biological pathways and networks related to stress induction were identified using Ingenuity Pathway Analysis (Ingenuity Systems, USA). Genes were selected as inputs for pathway testing when FDR was ≤0.05. Right-tailed Fisher’s exact test was conducted to calculate significance values of pathway fittings.
RESULTS Characteristics of participants Average participant age was 31.4 years (SD 11.6). Most study participants were Asian (90.0%), and ethnicity was not assessed. Average BMI was in the normal range with a mean of 23.16 (SD 4.69) kg/m2. While the sample tended to be well educated, average per capita income was low with a mean of $19,298.39 per year (SD 12,921.45), because many participants were graduate/undergraduate students. Both groups were quite similar for all demographic characteristics (Table 1).
Table 1: Study participant characteristics
Effects of TSST on psychological stress and physiological measures RM-ANCOVA results were as follows: interactions between the within-subjects factor (time) and the between-subjects factor (TSST) were significant, showing that participants who completed the TSST had significantly higher scores on SUDS (F1,32 = 59.89, p = 0.000, and η2 = 0.65) and state anxiety (F1, 35 = 10.62, p = 0.002, and η2 = 0.23) after stress induction compared to the control group, after controlling for age, MAP and BMI. Participants in the experimental group also showed significantly higher systolic BP (F1,35 = 26.53, p = 0.000, and η2 = 0.43), diastolic BP (F1,35 = 18.76, p = 0.000, and η2 = 0.34) and HR (F1,35 = 7.49, p = 0.010, and η2 = 0.18) after stress induction, compared with the control group, after controlling for age and BMI (Table 2).
Table 2: Trier Social Stress Test effects on psychological stress and physiological measures
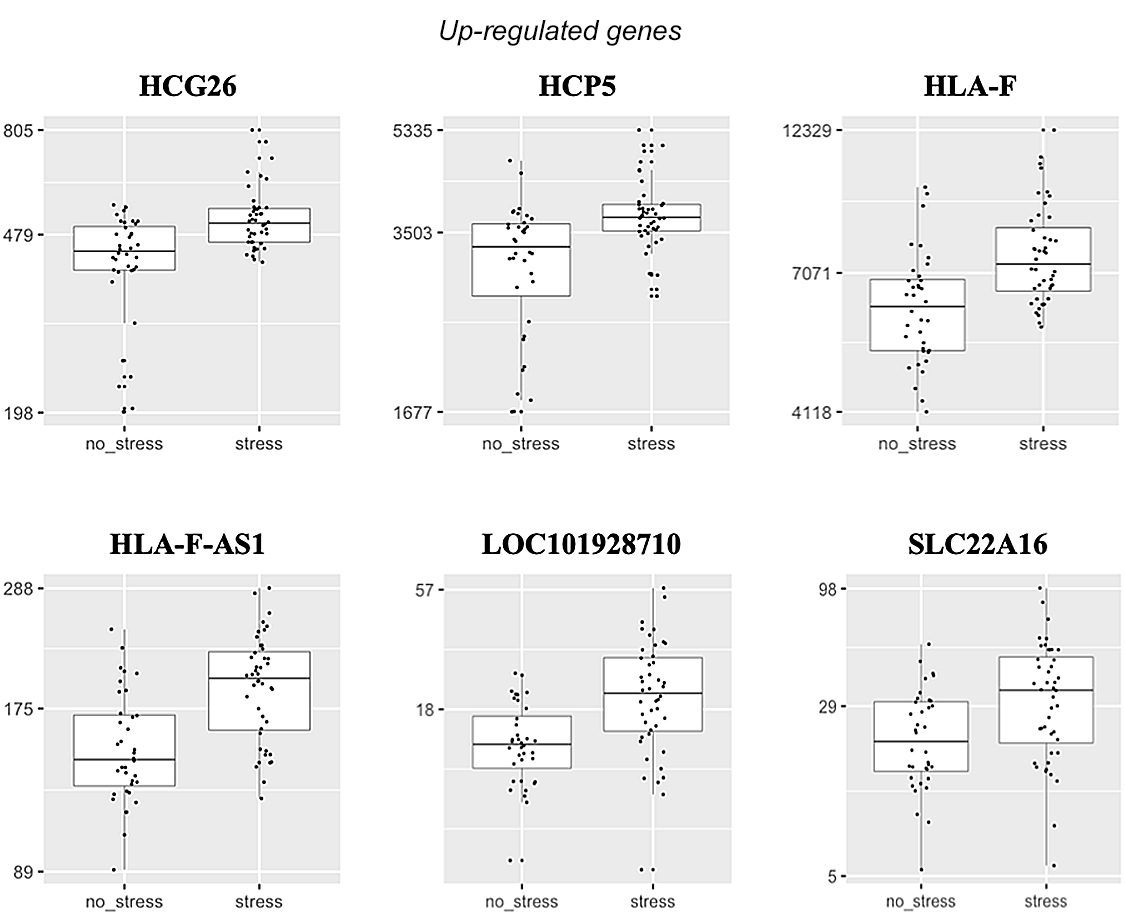
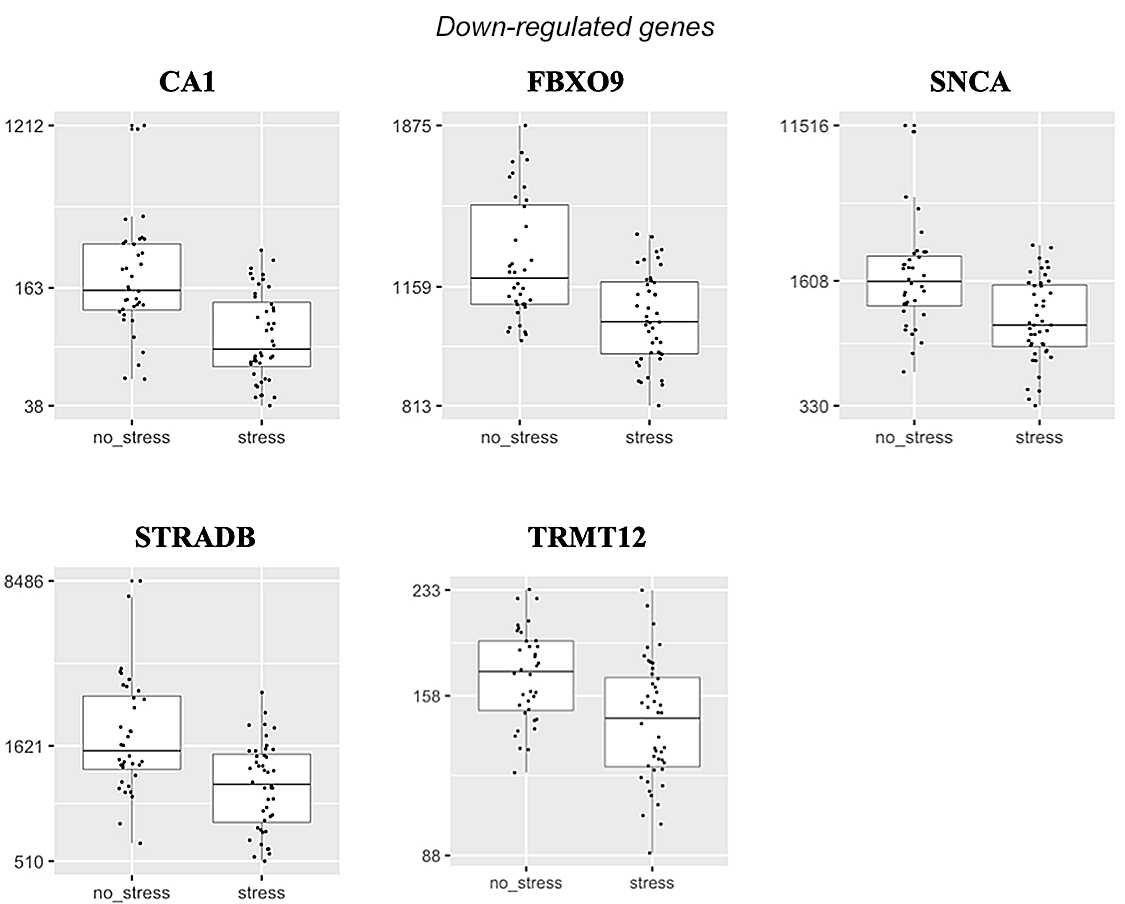
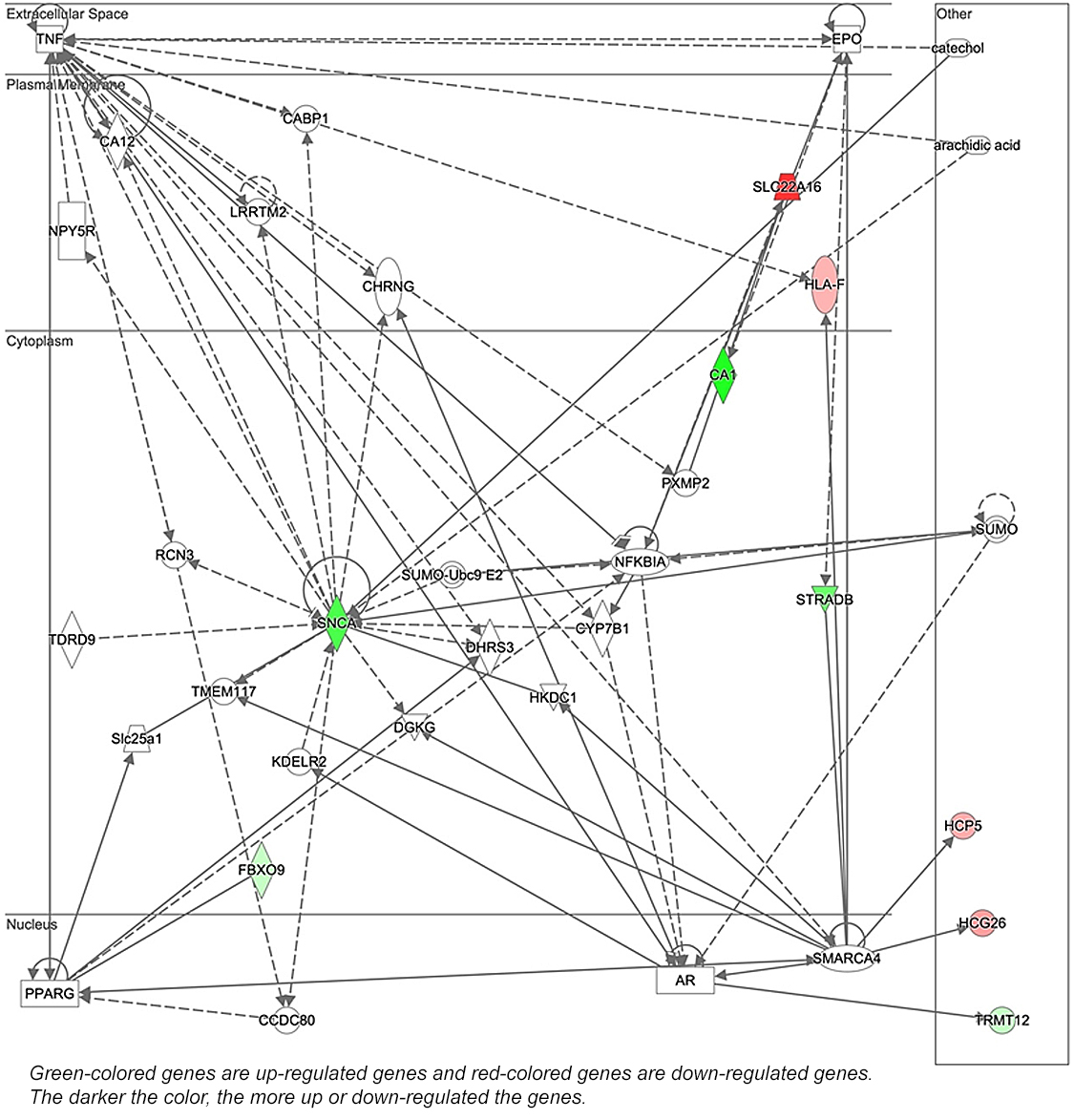
Differentially-expressed genes after stress induction in experimental group compared with control group Illumina’s NextSeq 500 sequencer generated around 30 million paired-end reads with read lengths of 75 bp (2 x 75 bp) per sample/library. A total of 22,021 genes were expressed. Tables 3 and 4 present the significantly up- and down-regulated genes associated with stress induction. Figure 1 shows normalized mRNA expression in terms of read counts for the differentially regulated genes under the acute stress condition (TSST). The Ingenuity Pathway Analysis (IPA) (QIAGEN, Germany) identified one related network; “Endocrine System Disorders, Neurological Disease, Organismal Injury and Abnormalities” with a score of 27 (Figure 2). Nine molecules of 11 were found in this network.
DISCUSSION This exploratory study investigated genes that were differently-expressed between two conditions that varied as to whether or not participants were exposed to the TSST, an acute psychological stressor. The psychological and physiological stress parameters (SUDS, state anxiety/STAI, BP and HR) indicated that the TSST successfully induced moderate—but statistically significant—levels of acute psychological stress in experimental group participants. Results showed significant up-regulation of six genes (HCG26, HCP5, HLA-F, HLA-F-AS1, LOC1019287, and SLC22A16) and down-regulation of five genes (CA1, FBXO9, SNCA, STRADB, and TRMT12) in the stress-induced group, compared with the control group.
Table 3: Up-regulated genes following stress induction in experimental group compared with control group
Table 4: Down-regulated genes after stress induction in experimental group compared with control group
IPA analysis demonstrated that 9 genes of 11 are implicated in endocrine system disorders, neurological disease, and organismal injury and abnormalities. This study revealed novel genes that have not been previously reported in relation to psychological stress. While this research was exploratory and requires confirmatory studies, it indicates that ultimately these genes may help elucidate pathophysiological mechanisms through which psychological stress is linked to disease conditions.
Up-regulation of genes linked to psychological stress induction Among six up-regulated genes, four genes (HCG26, HCP5, HLA-F, and HLA-F-AS1) are affiliated with the Human Leukocyte Antigen (HLA) complex, which is also referred to as the Major Histocompatibility Complex (MHC) in humans. The HLA complex is a group of proteins on the cell surface known to play a critical role in the immune system.[38] All four genes are also long non-coding RNAs (lncRNA), defined as RNA with more than 200 nucleotides that have no protein-coding capacity. The crucial function of lncRNA and its regulatory role in tumor occurrence and progression has been recognized in multiple studies.[39]
Histocompatibility leukocyte antigen complex p5 (HCP5) is known to play important roles in cancer cell proliferation, migration and invasion in many cancers.[40,41] With the down-regulation of HCP5’s anti-tumorigenic effect,[42] HCP5 is considered a promising biomarker and therapeutic target. On the other hand, only a few studies have been conducted on HCG26, HLA-F, and HLA-F-AS1, and the molecular mechanisms by which they are involved in biological processes remain largely unknown. HCG26 is in HLA complex group 26.
Figure 1: Normalized read counts (Y-axes) for differentially-expressed genes in experimental group (stress) compared with control group (no stress)
One study exploring the roles of lncRNAs in follicular development demonstrated that HCG26 expression was up-regulated in patients with polycystic ovary syndrome and was associated with follicle count and cell proliferation.[43] HLA-F encodes HLA Class I Histocompatibility Antigen, Alpha Chain F.[44] A previous study on patients with breast cancer showed that HLA-F expression was positively associated with tumor size and poor clinical outcomes.[45] HLA-F antisense RNA 1 (HLA-F-AS1) has been reported as up-regulated in colorectal cancer cell tissues and could promote colorectal cancer cell proliferation.[46] Acute stress induces leukocyte redistribution and increases cellular adhesion, molecule expression and chemotaxis, all of which are critical in immune cell recruitment and migration.[12] As per acute psychological stressor’s effects on leukocytosis, our findings suggest that acute psychological stress may dysregulate these four lncRNAs in the HLA system, which are known to play an important role in immune responses.
While we still do not know LOC1019287’s function, the function of SLC22A16 has been well-documented.[47] Membrane transporters are proteins that carry molecules across the cell membrane. Solute carrier (SLC) transporters are one of two large groups of membrane transporters. The SLC22 family contains cation and carnitine transporters, which include SLC22A16.[47] Previous studies have suggested that SLC22A16 may be a novel target for cancer treatment. For example, a study on acute myeloid leukemia demonstrated that SLC22A16 showed the greatest differential expression in acute myeloid leukemia cells among different carnitine transporters, compared with normal cells.[48]
Another study identified SLC22A16 as one of 13 hub genes involved in nasopharyngeal carcinoma carcinogenesis or progression, and could be useful as a diagnostic biomarker for nasopharyngeal carcinoma.[49] It has also been reported that SLC22A16 up-regulation is an independent unfavorable prognostic indicator in gastric cancer.[50] While the results from our exploratory analyses need to be interpreted cautiously, coupled with previous reports on the high correlation between SLC22A16 and different types of cancer, they can inform future studies that may elucidate the causal relationship between stress and cancer.
Down-regulation of genes linked to psychological stress induction SNCA has previously been associated with psychological stress in animal models.[51] SNCA encodes α-synuclein, one of three families of synuclein that are soluble proteins found in nervous system tissue. Variants within SNCA have been studied extensively due to SNCA’s link to Parkinson disease.[52] α-synuclein is known to have a role in synaptic vesicle cycling,[53] and overexpression of SNCA has a detrimental effect on neuron function.[52] Plasma and serum α-synuclein are suggested potential diagnostic biomarkers in Parkinson disease patients.[54]
There is interest in the association between psychological stress and Parkinson disease, and one study showed that chronic mild stress accelerates aggregation of α-synuclein in male mice,[51] suggesting a different directional effect than observed in this study. Given our result suggesting that acute stress down-regulates SNCA, it appears that acute and chronic psychological stress may exert different effects on SNCA expression, but clearly both the animal models and our study procedures need to be replicated and extended.
Down-regulated genes after stress induction also included CA1, F-box only protein 9 (FBXO9), STRADB, and TRMT12. CA1 encodes carbonic anhydrase 1 which belongs to a family of zinc metalloenzymes. CA1 is known to catalyze the reversible hydration of carbon dioxide and be involved in the regulation of hemoglobin’s affinity for oxygen.[55] A previous study on patients with colon cancer showed that higher CA1 expression levels were linked to higher survival probability than lower CA1 expression levels, and identified CA1 as a potential biomarker due to its predictive role in colon cancer status and survival time.[56] A recent study showed that CA1 is up-regulated in septic patients, suggesting the protective response of white cells in hostile environments like sepsis.[57] CA1 down-regulation in our study suggests that the gene may respond differently to severe stress conditions. FBXO9 is a member of the F-box protein family, which constitutes one of the four subunits of the ubiquitin protein ligase. A study showed that primary tumors with FBXO9 loss expressed high levels of proteins associated with metastasis and invasion.[58] STRADB encodes STE20-related kinase adapter protein beta enzyme, which is involved in cell cycles and apoptosis.[59] Reduced STRADB expression is associated with increased cell cycle length and consequent slowing down of the cell cycle.[60] TRMT12 is one of the tRNA methyltransferases that catalyze RNA methylation. TRMT12 is highly expressed in a large cohort of primary tumors,[61] and different cancer cell lines.[62]
Figure 2: The network identified by Ingenuity Pathway Analysis
Although not extensively studied, previous research suggests that CA1, FBXO9, and TRMT12 are implicated in cancer development. Future studies are needed to explore how various types of stress (e.g., acute vs. chronic psychological stress, or psychological vs. physiological stress) may differently regulate expression of these genes.
The strengths of this study include using a genomic approach to explore stress responses in healthy human subjects, use of RNA-sequencing, which is superior to the use of microarrays in gene expression profiling,[63] and successful induction of psychological stress in a laboratory setting, using the TSST. Nevertheless, this study has several limitations. First, relatively few genes were differentially expressed in the two groups.
Furthermore, inflammatory genes like cytokines, that have shown differential expression in prior studies, have not been expressed differently in the two groups of our study. This may be due to our study’s convenience sampling. Selecting participants that exhibited changes in arterial stiffness after stress induction may generate the results that reduce generalizability of the study findings, given that the participants may be more likely to demonstrate hemodynamic changes compared to others. Second, because this sample is small, and includes only females, the majority of whom were of Asian descent, sex, race and other confounders should also be considered. Third, while use of gene expression patterns in whole blood cells is convenient and may have advantages for translational research, RNA-seq studies of homogeneous cell populations or specific tissues can be more informative.[64] Fourth, people with high levels of trait anxiety are more likely to respond in stressful situations with increased anxiety. Therefore, future studies should explore whether higher scores on trait anxiety play a role in transcriptional activity related to acute stress situations. Fifth, the findings from this exploratory study should be replicated in a study conducted with a larger sample that includes quantitative reverse transcription PCR (RT-qPCR). Last, the potential confounding effects of lymphocyte subset redistribution were not controlled for in analyses. Lymphocyte redistribution in response to acute stress and sympathetic nervous system activation has been well-described;[65] thus, isolated leukocyte subpopulation analysis should be considered in future studies.
CONCLUSION This study highlights previously unreported associations of 11 genes with acute psychological stress and provides further evidence of stress-induced alterations of the gene expression profile. At present, we cannot explain the molecular mechanisms of these genes in stress responses. Future followup studies should validate the identified genes and explore the underlying mechanisms linking psychological stress, the identified genes, and their associated diseases, including cancer and neurodegenerative diseases. Such studies will clarify pathological mechanisms by which vulnerabilities to the diseases may be initiated or aggravated by psychological stress and may ultimately identify therapeutic targets that will enhance biological resilience to adverse effects of psychological stress.
FINANCING This study was supported by the K23NR016215 grant from the National Institutes of Health/National Institute of Nursing Research.
Iwata M, Ota KT, Duman RS. The inflammasome: pathways linking psychological stress, depression, and systemic illnesses. Brain Behav Immun. 2013 Jul;31:105–14. PMCID: PMC4426992.
Sillivan SE, Jones ME, Jamieson S, Rumbaugh G, Miller CA. Bioinformatic analysis of long-lasting transcriptional and translational changes in the basolateral amygdala following acute stress. PLoS One [Internet]. 2019 Jan 10 [cited 2021 Dec 10];14(1):e0209846. Available at: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0209846. PMID: 30629705.
Garner JP. The significance of meaning: why do over 90% of behavioral neuroscience results fail to translate to humans, and what can we do to fix it? ILAR J. 2014;55(3):438–56. PMID: 25541546.
McInnis CM, Wang D, Gianferante D, Hanlin L, Chen X, Thoma MV, et al. Response and habituation of pro- and anti-inflammatory gene expression to repeated acute stress. Brain Behav Immun. 2015 May;46:237–248. PMID: 25683696.
Huang CJ, Stewart JK, Franco RL, Evans RK, Lee ZP, Cruz TD, et al. LPS-stimulated tumor necrosis factor-alpha and interleukin-6 mRNA and cytokine responses following acute psychological stress. Psychoneuroendocrinology. 2011 Nov;36(10):1553–61. PMID: 21600698.
Kuebler U, Zuccarella-Hackl C, Arpagaus A, Wolf JM, Farahmand F, von Känel R, et al. Stress-induced modulation of NF-κB activation, inflammation-associated gene expression, and cytokine levels in blood of healthy men. Brain Behav Immun. 2015 May 1;46:87–95. PMID: 25557189.
Slavish DC, Szabo YZ. The effect of acute stress on salivary markers of inflammation: a systematic review protocol. Syst Rev. 2019 May 2;8(1):108. https://doi.org/10.1186/s13643-019-1026-4. PMID: 31046830; PMCID: PMC6498465.
Xiang L, Del Ben KS, Rehm KE, Marshall GD Jr. Effects of acute stress-induced immunomodulation on TH1/TH2 cytokine and catecholamine receptor expression in human peripheral blood cells. Neuropsychobiology. 2012;65(1):12–9. PMID: 22094268.
Zaba M, Kirmeier T, Ionescu IA, Wollweber B, Buell DR, Gall-Kleebach DJ, et al. Identification and characterization of HPA-axis reactivity endophenotypes in a cohort of female PTSD patients. Psychoneuroendocrinology. 2015 May;55:102–15. PMID: 25745955.
Redwine L, Snow S, Mills P, Irwin M. Acute psychological stress: effects on chemotaxis and cellular adhesion molecule expression. Psychosom Med. 2003 Jul–Aug;65(4):598–603. PMID: 12883110.
Nowotny B, Cavka M, Herder C, Löffler H, Poschen U, Joksimovic L, et al. Effects of acute psychological stress on glucose metabolism and subclinical inflammation in patients with post-traumatic stress disorder. Horm Metab Res. 2010 Sep;42(10):746–53. PMID: 20665427.
Schwaiger M, Grinberg M, Moser D, Zang JCS, Heinrichs M, Hengstler JG, et al. Altered stress-induced regulation of genes in monocytes in adults with a history of childhood adversity. Neuropsychopharmacology. 2016 Sep;41(10):2530–40. PMCID: PMC4987852.
Nater UM, Whistler T, Lonergan W, Mletzko T, Vernon SD, Heim C. Impact of acute psychosocial stress on peripheral blood gene expression pathways in healthy men. Biol Psychol. 2009 Oct;82(2):125–32. PMCID: PMC7116965.
Birkett MA. The Trier Social Stress Test protocol for inducing psychological stress. J Vis Exp. 2011 Oct 19;(56). PMCID: PMC3227197.
Het S, Rohleder N, Schoofs D, Kirschbaum C, Wolf OT. Neuroendocrine and psychometric evaluation of a placebo version of the “Trier Social Stress Test.” Psychoneuroendocrinology. 2009 Aug;34(7):1075–86. PMID: 19307062.
Logan JG, Teachman BA, Liu X, Farber CR, Liu Z, Annex BH. Acute psychological stress, autonomic function, and arterial stiffness among women. Int J Psychophysiol. 2020 Sep;155:219–26. PMID: 32619458.
Tanner BA. Validity of global physical and emotional SUDS. Appl Psychophysiol Biofeedback. 2012 Mar;37(1):31–4. PMID: 22038278.
Spielberger CD, Gorsuch RL, Lushene RE, Vagg PR, Jacobs GA, editors. Manual for the State-Trait Anxiety Inventory. California: Consulting Psychologists Press; 1983 Jan 1.
Spielberger CD, Vagg PR. Psychometric properties of the STAI: a reply to Ramanaiah, Franzen, and Schill. J Pers Assess. 1984 Feb;48(1):95–7. PMID: 6707862.
Maxwell MH, Waks AU, Schroth PC, Karam M, Dornfeld LP. Error in blood-pressure measurement due to incorrect cuff size in obese patients. Lancet. 1982 Jul 3;2(8288):33–6. PMID: 6123760.
van der Valk ES, Savas M, van Rossum EFC. Stress and obesity: are there more susceptible individuals? Curr Obes Rep. 2018 Jun;7(2):193–203. PMID: 29663153.
Laurent S, Cockcroft J, Van Bortel L, Boutouyrie P, Giannattasio C, Hayoz D, et al. Expert consensus document on arterial stiffness: methodological issues and clinical applications. Eur Heart J. 2006 Nov;27(21):2588–605. PMID: 17000623.
Berntson GG, Bigger JT Jr, Eckberg DL, Grossman P, Kaufmann PG, Malik M, et al. Heart rate variability: origins, methods, and interpretive caveats. Psychophysiology. 1997 Nov;34(6):623–48. PMID: 9401419.
Dickerson SS, Kemeny ME. Acute stressors and cortisol responses: a theoretical integration and synthesis of laboratory research. Psychol Bull. 2004 May;130(3):355–91. PMID: 15122924.
Kudielka BM, Buske-Kirschbaum A, Hellhammer DH, Kirschbaum C. HPA axis responses to laboratory psychosocial stress in healthy elderly adults, younger adults, and children: impact of age and gender. Psychoneuroendocrinology. 2004 Jan;29(1):83–98. PMID: 14575731.
Brody S, Preut R, Schommer K, Schürmeyer TH. A randomized controlled trial of high dose ascorbic acid for reduction of blood pressure, cortisol, and subjective responses to psychological stress. Psychopharmacology (Berl). 2002 Jan;159(3):319–24. PMID: 11862365.
Juster RP, Perna A, Marin MF, Sindi S, Lupien SJ. Timing is everything: anticipatory stress dynamics among cortisol and blood pressure reactivity and recovery in healthy adults. Stress. 2012 Nov;15(6):569–77. PMID: 22296506.
Polheber JP, Matchock RL. The presence of a dog attenuates cortisol and heart rate in the Trier Social Stress Test compared to human friends. J Behav Med. 2014 Oct;37(5):860–7. PMID: 24170391.
DeLuca DS, Levin JZ, Sivachenko A, Fennell T, Nazaire MD, Williams C, et al. RNA-SeQC: RNA-seq metrics for quality control and process optimization. Bioinformatics. 2012 Jun 1;28(11):1530–2. https://doi.org/10.1093/bioinformatics/bts196
Dobin A, Davis CA, Schlesinger F, Drenkow J, Zaleski C, Jha S, et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics. 2013 Jan 1;29(1):15–21. https://doi.org/10.1093/bioinformatics/bts635
Anders S, Pyl PT, Huber W. HTSeq–a Python framework to work with high-throughput sequencing data. Bioinformatics. 2015 Jan 15;31(2):166–9. https://doi.org/10.1093/bioinformatics/btu638
Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol [Internet]. 2014 [cited 2021 Dec 15];15(12):550. Available at: https://genomebiology.biomedcentral.com/articles/10.1186/s13059-014-0550-8. https://doi.org/10.1186/s13059-014-0550-8
Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J Royal Statistical Soc Series B (Methodological). 1995;57(1):289–300. https://doi.org/10.1111/j.2517-6161.1995.tb02031.x
Liu B, Shao Y, Fu R. Current research status of HLA in immune-related diseases. Immun Inflamm Dis [Internet]. 2021 Jun [cited 2021 Dec 15];9(2):340–50. PMID: 33657268. Epub 2021 Mar 3. https://doi.org/10.1002/iid3.416
Beermann J, Piccoli MT, Viereck J, Thum T. Non-coding RNAs in development and disease: background, mechanisms, and therapeutic approaches. Physiol Rev [Internet]. 2016 Oct [cited 2021 Dec 15];96(4):1297–325. https://doi.org/10.1152/physrev.00041.2015. PMID: 27535639.
Zou Y, Chen B. Long non-coding RNA HCP5 in cancer. Clin Chim Acta. 2021 Jan;512:33–9. PMID: 33245911.
Yin D, Lu X. Silencing of long non-coding RNA HCP5 inhibits proliferation, invasion, migration, and promotes apoptosis via regulation of miR-299-3p/SMAD5 axis in gastric cancer cells. Bioengineered. 2021 Dec;12(1):225–39. PMID: 33371778.
Wang K, Yu X, Tao B, Qu J. Downregulation of lncRNA HCP5 has inhibitory effects on gastric cancer cells by regulating DDX21 expression. Cytotechnology [Internet]. 2021 Feb [cited 2021 Dec 15];73(1):1–11. Available at: https://link.springer.com/article/10.1007/s10616-020-00429-0. PMCID: PMC7817730.
Liu YD, Li Y, Feng SX, Ye DS, Chen X, Zhou XY, et al. Long noncoding RNAs: Potential regulators involved in the pathogenesis of polycystic ovary syndrome. Endocrinology. 2017 Nov 1;158(11):3890–9. PMID: 28938484.
Kochan G, Escors D, Breckpot K, Guerrero-Setas D. Role of non-classical MHC class I molecules in cancer immunosuppression. Oncoimmunology [Internet]. 2013 Nov 1 [cited 2021 Dec 15];2(11):e26491. Available at: https://www.tandfonline.com/doi/abs/10.4161/onci.26491. PMID: 24482746.
Harada A, Ishigami S, Kijima Y, Nakajo A, Arigami T, Kurahara H, et al. Clinical implication of human leukocyte antigen (HLA)-F expression in breast cancer. Pathol Int [Internet]. 2015 Nov [cited 2021 Dec 15];65(11):569–74. Available at: https://onlinelibrary.wiley.com/doi/10.1111/pin.12343. PMID: 26332651.
Huang Y, Sun H, Ma X, Zeng Y, Pan Y, Yu D, et al. HLA-F-AS1/miR-330-3p/PFN1 axis promotes colorectal cancer progression. Life Sci. 2020 Aug 1;254:117180. PMID: 31863778. Epub 2019 Dec 18.
Wu Y, Hurren R, MacLean N, Gronda M, Jitkova Y, Sukhai MA, et al. Carnitine transporter CT2 (SLC22A16) is over-expressed in acute myeloid leukemia (AML) and target knockdown reduces growth and viability of AML cells. Apoptosis. 2015 Aug;20(8):1099–108. PMID: 25998464.
Zhang JZ, Wu ZH, Cheng Q. Screening and identification of key biomarkers in nasopharyngeal carcinoma. Medicine (Baltimore). 2019 Nov;98(48):e17997. PMID: 31770211.
Zhao W, Wang Y, Yue X. SLC22A16 upregulation is an independent unfavorable prognostic indicator in gastric cancer. Future Oncol. 2018 Sep;14(21):2139–48. PMID: 29698084.
Wu Q, Yang X, Zhang Y, Zhang L, Feng L. Chronic mild stress accelerates the progression of Parkinson’s disease in A53T α-synuclein transgenic mice. Exp Neurol. 2016 Nov;285(Pt A):61–71. PMID: 27637804.
Patel D, Bordoni B. Physiology, Synuclein. StatPearls. 2020. PMID: 31985951.
Villar-Piqué A, Lopes da Fonseca T, Outeiro TF. Structure, function and toxicity of alpha-synuclein: the Bermuda triangle in synucleinopathies. J Neurochem. 2016 Oct;139 Suppl 1:240–55. PMID: 26190401.
Chang CW, Yang SY, Yang CC, Chang CW, Wu YR. Plasma and serum alpha-synuclein as a biomarker of diagnosis in patients with Parkinson’s disease. Front Neurol [Internet]. 2020 Jan 21 [cited 2021 Dec 15];10:1388. Available at: https://www.frontiersin.org/articles/10.3389/fneur.2019.01388/full. PMID: 32038461.
Supuran CT. Structure and function of carbonic anhydrases. Biochem J. 2016 Jul 15:473(14):2023–32. PMID: 27407171.
Liu Z, Bai Y, Xie F, Miao F, Du F. Comprehensive analysis for identifying diagnostic and prognostic biomarkers in colon adenocarcinoma. DNA Cell Biol. 2020 Apr;39(4):599–614. PMID: 32031891. Epub 2020 Feb 7.
Leite GGF, Scicluna BP, van der Poll T, Salomão R. Genetic signature related to heme-hemoglobin metabolism pathway in sepsis secondary to pneumonia. NPJ Syst Biol Appl [Internet]. 2019 Aug 1 [cited 2021 Dec 17];5:26. PMID: 31396396. Available at: https://www.nature.com/articles/s41540-019-0105-4
Hynes-Smith RW, Swenson SA, Vahle H, Wittorf KJ, Caplan M, Amador C, et al. Loss of FBXO9 enhances proteasome activity and promotes aggressiveness in acute myeloid leukemia. Cancers (Basel) [Internet]. 2019 Nov 3 [cited 2021 Dec 17];11(11):1717. PMID: 31684170. Available at: https://www.mdpi.com/2072-6694/11/11/1717
Zhong DS, Sun LL, Dong LX. Molecular mechanisms of LKB1 induced cell cycle arrest. Thorac Cancer [Internet]. 2013 Aug [cited 2021 Dec 17];4(3):229–33. PMID: 28920233.
de Freitas RCC, Bortolin RH, Lopes MB, Tamborlin L, Meneguello L, Silbiger VN, et al. Modulation of miR-26a-5p and miR-15b-5p exosomal expression associated with clopidogrel-induced hepatotoxicity in HepG2 cells. Front Pharmacol [Internet]. 2017 Dec 12 [cited 2021 Dec 17];8:906. https://doi.org/10.3389/fphar.2017.00906. PMID: 29311920.
Manning M, Jiang Y, Wang R, Liu L, Rode S, Bonahoom M, et al. Pan-cancer analysis of RNA methyltransferases identifies FTSJ3 as a potential regulator of breast cancer progression. RNA Biol. 2020 Apr;17(4):474–86. PMID: 31957540. Epub 2020 Jan 19.
Rodríguez V, Chen Y, Elkahloun A, Dutra A, Pak E, Chandrasekharappa S. Chromosome 8 BAC array comparative genomic hybridization and expression analysis identify amplification and overexpression of TRMT12 in breast cancer. Genes Chromosomes Cancer [Internet]. 2007 Jul; 46(7):694–707. https://doi.org/10.1002/gcc.20454. PMID: 17440925.
Zhao S, Fung-Leung WP, Bittner A, Ngo K, Liu X. Comparison of RNA-Seq and microarray in transcriptome profiling of activated T cells. PLoS One [Internet]. 2014 Jan 16 [cited 2021 Dec 15];9(1):e78644. https://dx.plos.org/10.1371/journal.pone.0078644. PMCID: PMC3894192.
Wong L, Jiang K, Chen Y, Hennon T, Holmes L, Wallace CA, et al. Limits of peripheral blood mononuclear cells for gene expression-based biomarkers in juvenile idiopathic arthritis. Sci Rep. 2016 Jul 7;6:29477. PMCID: PMC4935846.
Dhabhar FS, Malarkey WB, Neri E, McEwen BS. Stress-induced redistribution of immune cells – from barracks to boulevards to battlefields: a tale of three hormones – Curt Richter Award Winner. Psychoneuroendocrinology. 2012 Sep;37(9):1345–68. PMCID: PMC3412918.
THE AUTHORS
Jeongok Gang Logan (Corresponding author: jl3zj@virginia.edu), registered nurse with a master’s degree and a doctorate in nursing. Associate professor, University of Virginia School of Nursing, Charlottesville, Virginia, USA. https://orcid.org/0000-0002-6599-6975
Sijung Yun, senior bioinformatics scientist with a master’s degree and a doctorate in genetics. Chief Science Officer of Predictive Care, Inc., Bethesda, Maryland, USA. https://orcid.org/0000-0003-1948-3690
Bethany Ann Teachman, clinical psychologist with a master’s degree and a doctorate in psychology, Professor, College and Graduate School of Arts & Sciences, University of Virginia, USA. https://orcid.org/0000-0002-9031-9343
Yongde Bao, analytical chemist with a master’s degree and a doctorate in analytical chemistry. Associate professor, Department of Microbiology, Immunology and Cancer Biology, School of Medicine, University of Virginia, USA. http://orcid.org/0000-0002-9031-9343
Emily Farber, research specialist with a master’s degree in animal science. Laboratory specialist, Center for Public Health Genomics, Department of Public Health Sciences, School of Medicine, University of Virginia, USA. https://orcid.org/0000-0003-3931-1781
Charles Ronnie Farber, human geneticist with a master’s degree and a doctorate in genetics. Associate professor and director, Center for Public Health Genomics, Department of Public Health Sciences, School of Medicine, University of Virginia, USA. https://orcid.org/0000-0002-6748-4711
No new Early Online articles at this time. The most recent articles are listed in the Current Issue Table of Contents. Early Online articles are added as soon as they are available, so please check back later.